









木质纤维素是一种来源广、含量丰富的可再生生物质资源,可以转化产生生物燃料、化工产品等多种产品,在农业、生物医药、工业及能源等领域具有重要的应用潜力[1-2]。木质纤维素由纤维素、半纤维素和木质素组成,通过共价和非共价键结合形成稳定的结构,使木质纤维素的降解成为制约其高效利用的一大瓶颈,大量富含木质纤维素的秸秆等农业废弃物被浪费[3-4]。酶法降解纤维素具有反应温和、环境污染小、催化活性稳定、转化率高和无毒性等优点,所利用的纤维素酶属于诱导型多组分复合酶系,主要包括外切葡聚糖酶、内切葡聚糖酶和β-葡萄糖苷酶,在降解纤维素过程中各组分相互协调发挥作用[5-6]。目前主要纤维素酶为丝状真菌分泌的胞外酶,但活性低且相对成本较高,制约了木质纤维素在酶解发酵等方面的应用。
越来越多的研究表明,存在多种其他类型的纤维素酶辅助纤维素降解过程。糖苷水解酶61家族(glycoside hydrolase 61, GH61)在真菌中广泛存在,例如生长在稻草、麦秆等富含纤维素基质上的草菇(Volvariella volvacea),其基因组中含有30个GH61家族同源基因[7]。研究发现GH61家族蛋白具有微弱的纤维素内切酶活性,但当和其他纤维素酶共同作用时,会使纤维素水解效率显著提高,表明其对纤维素酶类的水解具有促进作用,因此称为裂解多糖单加氧酶(lytic polysaccharide monooxygenase, LPMO),进一步被划分到辅助活性家族(auxiliary activity family protein, AA)[8-9]。黏质沙雷氏菌(Serratia marcesence)中的CBP21是第一个被发现的LPMO,后续大量作用于不同多糖底物的LPMO蛋白被发现,许多LPMOs并没有表现出严格的底物特异性,对半纤维素如木聚糖、木葡聚糖等底物也显示活性[10]。在CAZy数据库中,LPMOs现在被分为AA9(前身为GH61)、AA10(前身为CBM33)和AA11、AA13、AA14、AA15以及AA16,其中AA9优先作用于纤维素底物[11]。例如来自里氏木霉(Trichoderma reesei)的GH61家族的TrCel61A属于AA9家族的LPMO,在处理豆粕时可以显著提高内切葡聚糖酶和β-葡萄糖苷酶的糖化效率[12]。这类多糖单加氧酶(LPMOs)是一种金属依赖酶,2个反向平行的β-折叠面构成的结构可以结合Cu2+,在催化自然界中各种多糖的氧化裂解时发挥关键作用[13]。进一步对其作用方式分析发现,真菌GH61家族蛋白可以通过氧化纤维素链的C1-H或C4-H键生成糖醛酸等,导致纤维素糖苷链裂解并失去稳定状态、纤维素结构疏松,为其他纤维素酶提供作用位点,促进了其他纤维素酶发挥水解作用[14]。由于GH61蛋白在促进纤维素酶降解木质纤维素方面具有重要作用,因此开发稳定、高效辅助纤维素降解酶GH61,对于有效利用我国丰富的木质纤维素资源具有重要意义。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒
大斑刚毛座腔菌(CGMCC No9857),河北农业大学真菌毒素与植物分子病理学实验室分离,保存于中国微生物菌种保藏管理委员会普通微生物中心(CGMCC)。质粒pET32a载体购自生工生物工程(上海)股份有限公司,大肠杆菌Escherichia coli BL21(DE3)和Trans5α感受态细胞购自北京全式金生物技术(TransGen Biotech)股份有限公司。
1.1.2 培养基
1.1.3 试剂
质粒提取试剂盒购自天根生化科技(北京)有限公司,DNA胶回收试剂盒购自OMEGA公司,真菌RNA提取试剂盒购自上海生工公司,反转录试剂盒及染料法荧光定量预混试剂购自北京全式金公司,无缝克隆试剂盒购自北京博迈德基因技术有限公司,SDS-PAGE凝胶快速制备试剂盒购自赛国生物科技有限责任公司,限制性内切酶均购自宝日医生物技术(北京)有限公司,蛋白Marker购自北京索莱宝科技有限公司,纤维素酶购自上海麦克林生化科技股份有限公司(产品编号C6339-5g)。其他试剂均为国产分析纯。
1.2 方法
1.2.1 糖苷水解酶GH61家族氨基酸序列的获取及其生物信息学分析
在联合基因组研究中心JGI数据库(
对大斑刚毛座腔菌GH61家族蛋白进行以下分析,利用在线软件SignalP(
将大斑刚毛座腔菌GH61家族成员蛋白的氨基酸序列和番茄匍柄霉菌(Stemphylium lycopersici)、小麦褐斑长蠕孢霉菌(Pyrenophora triticirepentis)、稻平脐蠕孢菌(Bipolaris oryzae)、玉米小斑病菌(Bipolaris maydis)、细极链格孢菌(Alternaria tenuissima)、玉米圆斑病菌(Bipolaris zeicola)、链格孢叶斑病菌(Alternaria alternata)、麦根腐平脐蠕孢菌(Bipolaris sorokiniana)中的GH61家族蛋白进行系统发育分析,通过ClustalX进行序列比对后FastTree构建系统发育树,最后利用ITOL进行系统发育树的修饰。
1.2.2 大斑刚毛座腔菌的培养及滤纸酶活测定
将PDA培养基培养5 d的大斑刚毛座腔菌接种至CM液体基础培养基中25℃静置培养;将CM培养基的碳源葡萄糖替换为玉米秸秆粉末,同时接种大斑刚毛座腔菌25℃静置培养,以验证玉米秸秆对大斑刚毛座腔菌纤维素酶的诱导作用,上述处理设置3个重复。培养3、6和9 d后取上述两种培养基8 000 r/min离心10 min后的上清液,得到粗酶液。取1 cm×3 cm滤纸条放入20 mL具刻度试管中,加入0.5 mL粗酶液和1.5 mL柠檬酸-柠檬酸钠缓冲液,50℃恒温水浴50 min后取出,空白对照为酶液沸水浴处理10 min灭活,加入2 mL DNS试剂终止反应,摇匀后于沸水浴中处理5 min,取出冷却,利用分光光度计在540 nm处测定其吸光度值(OD值)并计算酶活力。定义在检测条件下,每分钟降解纤维素产生1 μmol葡萄糖所需要的酶量,称为一个酶活力单位(U)。
1.2.3 转录组测定GH61家族基因的转录水平
对上述两种培养基中培养7 d的大斑刚毛座腔菌的菌丝进行转录组测序。过滤菌丝置于液氮中,用于提取RNA。将样品利用高通量组织研磨仪液氮研磨,使用真菌RNA提取试剂盒并按照说明书操作提取RNA。将提取得到的RNA反转录cDNA。样品总RNA经检测合格后送至美吉生物公司进行RNA-Seq测序及分析。以葡萄糖为碳源培养的菌丝样品编号为“glucose”,以玉米秸秆为碳源培养的菌丝样品编号为“corn stover”。筛选差异倍数FC(Fold Change)>2且P<0.05的GH61家族基因,并利用R语言包绘制其基因表达水平的热图,用于鉴定参与木质纤维素利用的GH61家族蛋白。
1.2.4 实时荧光定量PCR检测GH61基因的表达
将在上述两种不同碳源下培养6 d和9 d的大斑刚毛座腔菌菌丝提取RNA并反转录cDNA。利用实时荧光定量PCR验证转录组分析筛选得到的GH61基因的相对表达水平。比对GH61家族基因获得非保守区域用于设计引物,并使用β-微管蛋白(β-tubulin)基因作为内参基因(表1)。使用Trans Start Green qPCR Super Mix试剂配置体系并设置反应程序,操作方法参照试剂说明书。使用CFX ConnectTM(Bio-Red,美国)进行RT-qPCR扩增及检测,利用 2-ΔΔCT法进行数据处理,计算各基因的相对表达量。
表1 实时荧光定量PCR引物序列
Table 1
| 引物名称Primer name | 序列Sequence(5'-3') | 引物名称Primer name | 序列Sequence(5'-3') | |
|---|---|---|---|---|
| StGH61_1-F | GGCTCTGGAACTGGCAAGAT | StGH61_12-F | CTCCTGGTCCGTCACTGTTC | |
| StGH61_1-R | TAGTACTCGAGGTCCCTGGC | StGH61_12-R | GGCTGGAGAGATCGGTCATG | |
| StGH61_2-F | GTCAGTTTCCCGGGTGCTTA | StGH61_13-F | GGCAGAAGGACCAGATCGAG | |
| StGH61_2-R | GACCACGCTGTTAGTCCCAA | StGH61_13-R | CGCTGTAGAGACCGGGAATC | |
| StGH61_3-F | ACATCAACACGTGGGATCCC | StGH61_14-F | CGACGCCATCCTAGACACTC | |
| StGH61_3-R | TGCGCACGACAGGTAGAATT | StGH61_14-R | TTGAAGTCGATGCCTGGGTC | |
| StGH61_4-F | CGACTTCCGTTGCAACAAGG | StGH61_15-F | CACCAAAGTCGAGCCCTTCT | |
| StGH61_4-R | AACATCACCAGGAGCCTTGG | StGH61_15-R | TTGAGGGAGAGGGAGACGAG | |
| StGH61_5-F | TTGCGCAAATCACAACCTCG | StGH61_16-F | GTGCCAAGGGAGGTCTCTTC | |
| StGH61_5-R | TCGTGGCATGTCATGTTGGA | StGH61_16-R | GGTCGGCAGTGAAAGAGACA | |
| StGH61_6-F | CACCCAGACTGTCACCATCC | StGH61_17-F | GGTTCAAGGTGCAGGAGGAA | |
| StGH61_6-R | CACCCAGACTGTCACCATCC | StGH61_17-R | CTACCCGTCACCTTGAGCTG | |
| StGH61_7-F | GGTCCAAGTTCTCGCAGGAA | StGH61_18-F | AGCTCGACTGCCATGATCTG | |
| StGH61_7-R | CATTCACATTCGAGCCGCTG | StGH61_18-R | GGGGAGCTTGACGTTGATGA | |
| StGH61_8-F | CCACAAGAATGCTAGCCCCA | StGH61_19-F | CAAGGTCTCCAACGCAGCTA | |
| StGH61_8-R | GCACCCTCAATCTTGGTCCA | StGH61_19-R | AAAATTGAGCACCGCCAACC | |
| StGH61_9-F | TGGTTCAAGGTTTCCAGCGA | StGH61_20-F | CATCCCTGCTTGTATTGCGC | |
| StGH61_9-R | AAATGTAGAATTGCGCGCCG | StGH61_20-R | GGAACGAGACGGTTGATGGT | |
| StGH61_10-F | GGGCAGTGATGTGAAGAGCT | StGH61_21-F | AAGATCGACGAGCAAGGCAT | |
| StGH61_10-R | GGAGCACGCGAGGTAGAATT | StGH61_21-R | GACGCATACCCAGGAATCGT | |
| StGH61_11-F | AGCCTCTCTTCCTCGGACAT | β-tubulin-F | GTGCGCAAGGAGGCTGAGGG | |
| StGH61_11-R | TTTTGGCGTCGCTGACTTTG | β-tubulin-R | CATGAAGAAATGGAGACGGGGGAA |
1.2.5 StGH61-11重组质粒的构建
以大斑刚毛座腔菌cDNA为模板,利用引物S1/S2(5'-CTGATATCGGATCCGAATTCATGTCGACATGGCAGCAACT-3'/5'-TGGTGGTGGTGGTGCTCGAGACCAGAGTACACAGCGGGTC-3')扩增StGH61-11片段。PCR反应条件为95℃ 3 min ;94℃ 30 s,55℃ 30 s,72℃ 5 min,共34个循环;72℃ 10 min。PCR产物经胶回收试剂盒回收片段后与Xho I和EcoR I酶切后的pET32a(+)进行连接,连接产物转化到大肠杆菌Trans5α中,在含有氨苄抗性的LB平板上37℃过夜培养,筛选阳性克隆送北京华大公司测序。经测序正确后得到重组质粒pET32a-StGH61-11。
1.2.6 StGH61-11重组蛋白的诱导表达及条件优化
将重组质粒转化大肠杆菌BL21,氨苄抗性进行PCR筛选阳性转化子,将阳性转化子接种于含50 μg/mL氨苄抗性的LB培养基中,37℃ 220 r/min振摇至菌体OD600为0.6-0.8后,加入0.2-1 mmol/L的IPTG,25℃,200-220 r/min摇床培养0-15 h,诱导蛋白表达。利用12% SDS-PAGE检测诱导蛋白含量,并使用2,6-DMP法进行StGH61-11酶活力测定,得到最佳诱导条件[19]。
1.2.7 2,6-DMP法测定StGH61-11重组蛋白LPMO酶活
将860 μL 116 mmol/L pH值为6.0的磷酸盐缓冲液,100 μL 100 mmol/L 2,6二甲氧基苯酚溶液(2,6-DMP)和20 μL 5 mmol/L H2O2储备溶液充分混合,孵育15 min。再加入20 μL的重组蛋白StGH61-11,与反应液混合均匀,30℃反应5 min,在469 nm处测定吸光值。酶活力定义为在反应条件下,每分钟转化2 μmo1 2,6-DMP形成1 μmo1四甲氧联对苯醌定义为1个LPMO酶活力单位。
1.2.8 温度、pH对StGH61-11重组蛋白酶活的影响
为了测定最适反应温度,取1 mL粗酶液,CMC-Na用pH 4.8的柠檬酸-柠檬酸钠缓冲液溶解,配置成1%的溶液,将反应体系放入30、40、50和60℃水浴锅中反应30 min,DNS法检测还原糖,计算重组酶在不同温度下的活力,并以最高酶活为100%,计算其他条件下的相对酶活。为探究其温度稳定性,将重组酶分别置于30、50和60℃水浴中,于3、6和9 h取出,分别测定酶活力,并以最高酶活为100%,计算其他条件下的相对酶活。
为了测定最适反应体系的pH,将重组酶与不同pH的底物于最适温度下反应30 min。所使用的缓冲体系pH为4、5、6和7,0.1 mol/L柠檬酸-柠檬酸钠缓冲液;pH 8、9、10和11,0.1 mol/L Tris-盐酸缓冲液。以最高酶活为100%,计算其他条件下的相对酶活。为了探究其pH值稳定性,将重组酶分别置于不同pH值的缓冲液中,在3、6和9 h取出,分别测定酶活力,并以最高酶活为100%,计算其他条件下的相对酶活。
1.2.9 StGH61-11蛋白与纤维素酶对木质纤维素水解的协同效应
以玉米秸秆为底物,将StGH61-11蛋白与纤维素酶对玉米秸秆进行协同酶解反应。在pH 5的PBS缓冲液体系中处理5%的玉米秸秆,加入5 U/mL的纤维素酶,并分别添加0.4、0.8和1.2 mg/mL重组蛋白StGH61-11。将上述各反应体系在50℃条件下反应6、12、24和48 h。以分别单独添加0.4 mg/mL重组蛋白StGH61-11和5 U/mL纤维素酶为对照。将反应后的体系进行离心,取上清液采用DNS法测定还原糖含量,计算糖化率(糖化率/%=总还原糖含量(mg)/总生物质质量(mg)×100%),并计算StGH61-11蛋白与纤维素酶作用的协同度(协同度=还原糖(StGH61-11+纤维素酶)/(还原糖StGH61-11+还原糖纤维素酶))。
2 结果
2.1 大斑刚毛座腔菌糖苷水解酶GH61家族蛋白特性及结构域
对大斑刚毛座腔菌基因组数据库进行搜索,获得21个GH61家族蛋白。对其蛋白性质进行生物信息学分析(表2),结果显示,大斑刚毛座腔菌GH61蛋白的氨基酸数目为149-472 aa,相对分子质量15.76-49.64 kD,等电点(pI)为5.47-9.00,除StGH61-14编码的蛋白为疏水性蛋白外(GRAVY值为0.024),其余蛋白均为亲水性蛋白。大多数GH61家族蛋白氨基酸序列的N端具有信号肽,但StGH61-9编码的蛋白没有信号肽切割位点。大部分蛋白分泌到胞外,而较少蛋白可以结合到细胞器的质膜上。
表2 大斑刚毛座腔菌糖苷水解酶GH61家族蛋白性质预测
Table 2
| 基因 Gene | 检索号 Accession number | 氨基酸数目 Number of amino acids | 分子量 Molecular weight/kD | 等电点 pI | 信号肽长度 Signal peptide length/aa | 蛋白定位 Protein location |
|---|---|---|---|---|---|---|
| StGH61-1 | XP_008027730 | 250 | 26.69 | 8.26 | 16 | 胞外Extracellular |
| StGH61-2 | XP_008031219 | 334 | 36.13 | 5.77 | 18 | 胞外Extracellular |
| StGH61-3 | XP_008024527 | 221 | 23.46 | 8.33 | 17 | 胞外Extracellular |
| StGH61-4 | XP_008025833 | 418 | 43.38 | 7.63 | 17 | 胞外Extracellular |
| StGH61-5 | XP_008021808 | 266 | 28.81 | 8.24 | 21 | 胞外Extracellular |
| StGH61-6 | XP_008023212 | 291 | 32.06 | 6.16 | 22 | 膜结合溶酶体Lysosomes |
| StGH61-7 | XP_008024957 | 248 | 26.32 | 7.69 | 20 | 胞外Extracellular |
| StGH61-8 | XP_008025253 | 329 | 34.56 | 5.98 | 22 | 分泌到胞外 Extracellular(Secreted) |
| StGH61-9 | XP_008030985 | 149 | 15.76 | 7.83 | — | 分泌到胞外 Extracellular(Secreted) |
| StGH61-10 | XP_008022828 | 236 | 25.55 | 9.00 | 20 | 膜结合线粒体Mitochondria |
| StGH61-11 | XP_008023697 | 229 | 23.52 | 8.49 | 16 | 胞外Extracellular |
| StGH61-12 | XP_008023696 | 258 | 27.49 | 5.47 | 22 | 胞外Extracellular |
| StGH61-13 | XP_008028494 | 325 | 34.02 | 6.31 | 18 | 胞外Extracellular |
| StGH61-14 | XP_008028319 | 292 | 30.73 | 6.28 | 21 | 胞外Extracellular |
| StGH61-15 | XP_008025489 | 472 | 49.64 | 7.44 | 19 | 胞外Extracellular |
| StGH61-16 | XP_008027303 | 222 | 23.26 | 7.69 | 20 | 胞外Extracellular |
| StGH61-17 | XP_008027295 | 246 | 25.92 | 8.44 | 18 | 胞外Extracellular |
| StGH61-18 | XP_008024355 | 344 | 34.75 | 6.49 | 17 | 胞外Extracellular |
| StGH61-19 | XP_008022590 | 243 | 25.06 | 8.90 | 17 | 胞外Extracellular |
| StGH61-20 | XP_008026441 | 233 | 24.21 | 7.67 | 18 | 胞外Extracellular |
图1
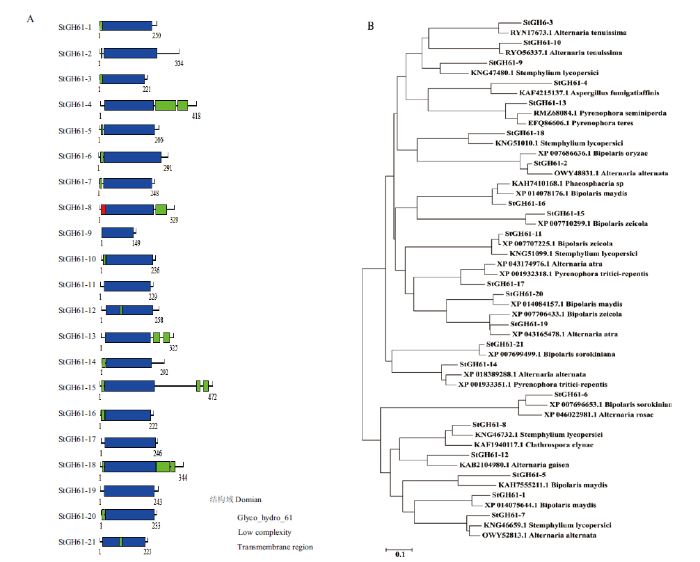
图1
大斑刚毛座腔菌糖苷水解酶GH61家族蛋白结构域及进化分析
A:大斑刚毛座腔菌糖苷水解酶GH61家族蛋白结构域;B:大斑刚毛座腔菌糖苷水解酶GH61家族蛋白进化关系
Fig. 1
Protein domains and evolutionary analysis of the GH61 family of glycoside hydrolases from S. turcica
A: Domain of GH61 family of glycoside hydrolases from S. turcica. B: Evolutionary relationship of glycoside hydrolases GH61 family proteins from S. turcica
2.2 玉米秸秆诱导下玉米毛座腔菌GH61家族基因的表达情况
在碳源分别为玉米秸秆和葡萄糖的培养基中培养大斑刚毛座腔菌,测定纤维素酶活性。结果(图2-A)显示,在培养基碳源为玉米秸秆时,纤维素酶活显著提高,当碳源为葡萄糖时大斑刚毛座腔菌培养9 d后滤纸酶活为4.5 U/mL,而碳源为玉米秸秆时滤纸酶活为27.5 U/mL,是葡萄糖为碳源时的6.11倍,说明此时大斑刚毛座腔菌分泌更多纤维素酶参与降解玉米秸秆中的木质纤维素。利用转录组分析两种培养条件下GH61家族基因的表达情况发现,GH61家族大多数基因表达上调(图2-B)。当限制条件为差异倍数<0.5且P<0.05时,GH61家族中的11个基因在玉米秸秆刺激下表达量显著升高,包括StGH61-3、StGH61-4、StGH61-7、StGH61-8、StGH61-11、StGH61-13、StGH61-14、StGH61-16、StGH61-18、StGH61-19、StGH61-20,其中StGH61-14表达量升高3.81倍、StGH61-7表达量升高5.75倍,StGH61-11表达量增加最多,升高6.54倍。为了验证转录组结果,利用荧光实时定量PCR检测了GH61家族21个基因的相对表达量,结果(图2-C)表明,StGH61-3、StGH61-11、StGH61-13、StGH61-14等多个基因表达水平在以玉米秸秆为碳源时显著增加,与转录组结果一致。
图2
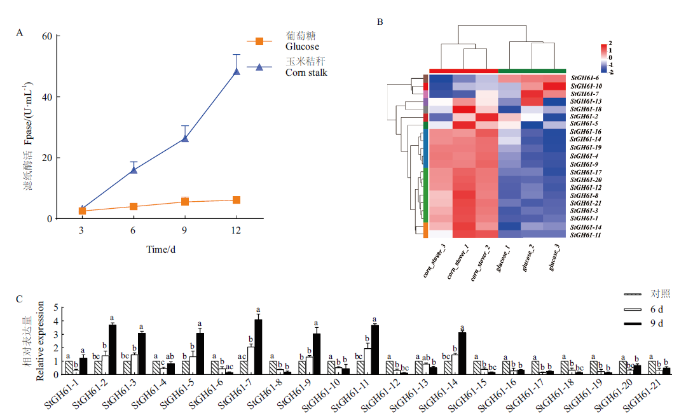
图2
大斑刚毛座腔菌纤维素酶的滤纸酶活及GH61家族基因表达分析
A:大斑刚毛座腔菌纤维素酶的滤纸酶活;B:大斑刚毛座腔菌中GH61家族糖苷水解酶基因的表达图谱;C:RT-qPCR验证大斑刚毛座腔菌中GH61家族糖苷水解酶基因相对表达量。不同小写字母表示同一基因在不同处理下存在显著差异(P<0.05)
Fig. 2
Filter paper enzymatic activity and GH61 family gene expression analysis of cellulase from S. turcica
A: Filter paper enzymatic activity of cellulase from S. turcica. B: Expression map of GH61 family glycoside hydrolase genes in S. turcica. C: RT-qPCR verifies the relative expression of GH61 family glycoside hydrolase genes in S. turcica. Different normal letters indicate significant differences among different treatments at 0.05 level
2.3 StGH61-11重组蛋白异源表达及诱导条件优化
选择在玉米秸秆诱导下表达水平增加最高的StGH61-11基因,进行异源表达。扩增StGH61-11的cDNA片段(图3-A),构建重组质粒PET32a-StGH61-11转化到大肠杆菌BL21中,PCR验证阳性转化子(图3-B)。利用IPTG诱导筛选得到的阳性转化子,蛋白表达后在细胞破碎上清液中检测到分子量约为42.6 kD的单一蛋白条带,与带有标签的StGH61-11重组蛋白预期大小相符(图3-C)。对异源表达蛋白诱导条件进行优化,在不同诱导时间对StGH61-11进行诱导表达并测定酶活,结果显示在诱导时间为9 h时酶活较高(图3-D)。用不同终浓度的IPTG对StGH61-11进行诱导表达,结果显示当终浓度为1.0 mmol/L时诱导酶活较高(图3-E),LPMO酶活可达到(54.08±1.67)U/g。
图3
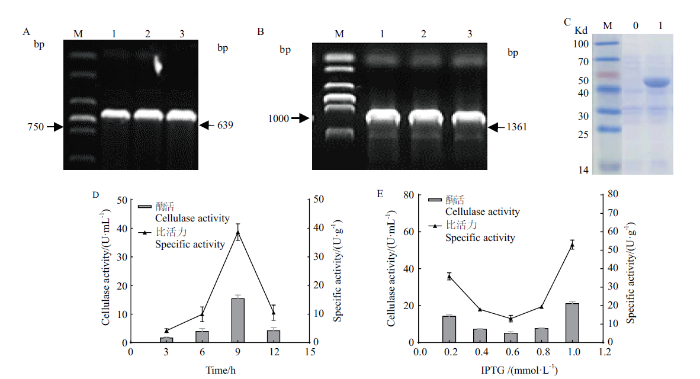
图3
StGH61-11重组蛋白异源表达及优化诱导条件
A:目的片段StGH61-11的PCR扩增产物琼脂糖凝胶电泳结果, 1-3:StGH61-11的cDNA片段, M:DNA marker;B:PCR验证重组转化子的琼脂糖凝胶电泳结果, 1-3:Pet32a-StGH61-11 重组质粒, M:DNA marker;C:SDS-PAGE 检测pET32a-StGH61-11(BL21)重组菌表达蛋白, 0:Pet32a-StGH61-11-BL21(DE3)重组菌未诱导对照, 1 :Pet32a-StGH61-11-BL21(DE3)重组菌经IPTG诱导后, M :预染蛋白分子量标准;D:诱导时间对重组蛋白StGH61-11酶活的影响;E:IPTG浓度对重组蛋白StGH61-11酶活的影响
Fig. 3
Heterologous expression of StGH61-11 recombinant protein and optimized induction conditions
A: Agarose gel electrophoresis results of the target gene StGH61-11 PCR products, 1-3: GH61-11 cDNA fragment. M:DNA marker. B: Agarose gel electrophoresis results of PCR for recombinant transformants, 1-3: Pet32a-StGH61-11 recombinant plasmid, M:DNA marker. C: SDS-PAGE detection of whole bacterial lysate expressed by recombinant pET32a-StGH61-11(BL21), 0 : Pet32a-StGH61-11-BL21(DE3)not induce control, 1 : Pet32a-StGH61-11-BL21(DE3)the recombinant bacteria were induced by IPTG, M : Standard for molecular weight of pre-dyed protein. D: Effect of induction time on the enzymatic activity of recombinant protein StGH61-11. E: Effect of IPTG concentration on the enzymatic activity of recombinant protein StGH61-11
2.4 StGH61-11的酶学性质分析
图4
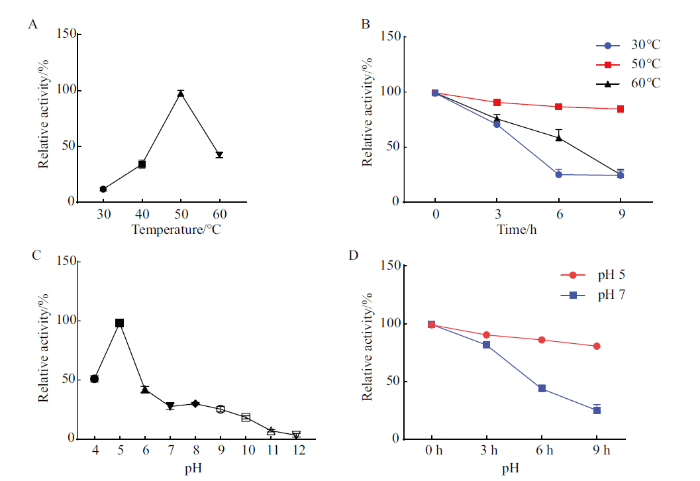
图4
StGH61-11重组蛋白的最适反应温度和稳定性及pH和稳定性
A:温度对StGH61-11纤维素酶活力的影响;B:温度对StGH61-11纤维素酶活力的稳定性影响;C:pH对StGH61-11纤维素酶活力的影响;D:pH对StGH61-11纤维素酶活力稳定性的影响
Fig. 4
Optimal reaction temperature and stability and pH and stability of StGH61-11 recombinant protein
A: Effects of temperature on the cellulase activity of StGH61-11. B: Effect of temperature on the stability of StGH61-11 cellulase activity. C: Effects of pH on StGH61-11 cellulase activity. D: Effect of pH on the viability stability of StGH61-11 cellulase
2.5 StGH61-11与纤维素酶在水解木质纤维素中的协同效应
为了检测重组蛋白StGH61-11对纤维素酶的促进作用,使用重组蛋白与5 U/mL的纤维素酶在最佳反应条件下共同处理5%玉米秸秆天然生物质原料,计算不同的酶体系在不同原料中的糖转化率和降解纤维素的协同度。结果(图5)显示,添加StGH61-11后体系中被水解的还原糖较未添加时明显增加,当1.2 mg/mL StGH61-11蛋白和纤维素酶协同处理玉米秸秆,反应12 h后体系中的还原糖含量较未添加时增加125%,糖化率达到46.5%,协同度达到2.50。表明添加StGH61-11能够显著地增强纤维素酶对玉米秸秆的水解作用。
图5

图5
StGH61-11 与纤维素酶水解玉米秸秆的协同反应
A:StGH61-11对纤维素酶活性的促进作用;B:StGH61-11与纤维素酶共同作用的协同度
Fig. 5
Synergistic reaction of StGH61-11 with cellulase hydrolysis of corn stover
A: Promoting effect of StGH61-11 on cellulase activity. B: Synergism degree of StGH61-11 and cellulase
3 讨论
3.1 大斑刚毛座腔菌GH61家族基因表达受到木质纤维素底物的诱导
糖苷水解酶GH61分布非常广泛,绝大多数来源于真菌,而且在基因组中以多基因家族形式存在。在米曲霉(Aspergillus oryzae)中分析发现了10种GH61家族基因[20];桧状青霉(Penicillium piceum)具有较为均衡的纤维素水解酶系,其基因组中含有1个GH61家族基因[21]。草菇(Volvariella volvacea)中有30个GH61家族基因,其编码蛋白分子量约22.4-100.2 kD,理论等电点为5.2-9.3[7]。本文在大斑刚毛座腔菌中发现21个编码GH61蛋白的基因,均具有GH61家族蛋白保守结构域,与番茄匍柄霉菌、玉米圆斑病菌及链格孢菌属真菌等多种植物病原真菌的GH61蛋白同源。但大斑刚毛座腔菌GH61家族编码蛋白的氨基酸数目为149-472 aa,差距较大,说明其功能可能存在差异。
研究发现木质纤维素可以诱导真菌纤维素水解酶酶活增加,同时GH61家族基因表达水平增加。通过对以甘蔗渣作为唯一碳源的培养基上培养的塔马里曲霉(Aspergillus tamarii)转录组分析发现,GH61家族中的5个基因表达水平显著上调[22]。本研究同样发现大斑刚毛座腔菌在玉米秸秆为碳源时的纤维素滤纸酶活是以葡萄糖为碳源培养时的6.11倍,表明木质纤维素对大斑刚毛座腔菌纤维素水解相关酶类的表达具有诱导作用。进一步利用转录组及实时荧光定量PCR验证筛选得到StGH61-2、StGH61-7、StGH61-14等多个GH61家族基因在降解秸秆过程中均上调,其中StGH61-11表达量上调最明显,与葡萄糖培养时相比升高6.54倍。因此本文以StGH61-11为进一步研究对象,异源表达并研究其对纤维素降解的促进作用。
3.2 LPMO蛋白StGH61-11可以促进纤维酶水解木质纤维素
本研究成功诱导表达蛋白StGH61-11并优化其诱导条件,在诱导9 h时,诱导StGH61-11酶活最高,实现了在短时间诱导条件下得到更高的酶活,LPMO酶活可达到(54.08±1.67)U/g,但重组蛋白只显示出微弱的纤维素酶活(25 U/mL)。烟曲霉(A. fumigatus)中AfAA9_B在大肠杆菌中异源表达后活力可达(8.33±1.9)U/g,同时对纤维素酶水解效率的促进作用达到20%左右;在大肠杆菌中异源表达来源于嗜热毁丝霉(Myceliophthora thermophila)的MtC1,其LPMO酶活力可达10.3 U/g[23-24]。由此可见本研究获得了具有较高LPMO酶活的AA9重组蛋白。对其酶学性质分析发现,温度和pH对StGH61-11酶活力具有明显影响,最适温度和pH值分别为50℃和5.0,同时在温度为50℃和pH值5.0时,处理9 h,依然能保持90%以上的活力,表明StGH61-11的稳定性良好。对其他真菌的GH61家族糖苷水解酶研究发现,最适温度在40-60℃,最适pH值偏酸性,在4.0-6.0左右[25⇓-27]。StGH61-11与之前对其他真菌GH61酶活的研究相似,在温度较高且偏酸性环境中活性较好,同时与其他GH61蛋白相比其温度及pH稳定性较高,具有应用潜力。LPMO对温度的稳定性分析显示,StGH61-11在30℃反而比在50和60℃更不稳定,这种现象可能与不同温度对催化底物H2O2的影响及蛋白构象的影响有关,具体原因需要进一步研究[28-29]。
当LPMO蛋白与其他纤维素酶共同作用时会促进其纤维素水解效率,对其进行深入研究是提高木质纤维素降解的另一种有效途径。通过异源表达黑曲霉(Asperillus niger)筛选得到的GH61家族基因发现,其与纤维素酶协同作用于玉米芯和滤纸时,水解得到的还原糖分别比纤维素酶单独作用时高24.69%和46.61%,协同度为1.086-1.288,促进了纤维素酶对底物的水解作用[30]。本文研究发现StGH61-11也能够显著地增强纤维素酶对木质纤维素的水解作用,并且StGH61-11的浓度越高,协同效应越显著。当添加StGH61-11蛋白1.2 mg/mL反应12 h,还原糖产量较未添加时增加125%,糖化率可达46.5%,协同度达到2.50,表明异源表达的StGH61-11促进了木质纤维素的水解,具有应用价值。本文通过表征木质纤维素底物对基因表达水平,筛选发挥作用的关键基因并对其进行高效表达,获得了具有促进纤维素酶降解木质纤维素的重组蛋白StGH61-11,为进一步高效降解木质纤维素的应用提供依据。
4 结论
木质纤维素玉米秸秆诱导下大斑刚毛座腔菌纤维素滤纸酶活增加6.11倍,且21个GH61家族基因中StGH61-11等11个基因的相对表达水平明显升高。在1.0 mmol/L IPTG 25℃诱导9 h后获得的重组蛋白StGH61-11,酶活可达(54.08±1.67)U/g。重组蛋白最适反应温度为50℃,最适反应pH为5.0,且在最适条件下保持9 h酶活仍维持90%以上。重组StGH61-11可以促进纤维素酶降解玉米秸秆,协同度最高达2.50,糖化率最高达46.5%,在酶解木质纤维素中具有应用价值。
参考文献
Characterization of a GH8 β-1, 4-glucanase from Bacillus subtilis B111 and its saccharification potential for agricultural straws
[J].DOI:10.4014/jmb.2105.05026 URL [本文引用: 2]
木质纤维素的微生物降解
[J].
Microbial degradation of lignocellulose
[J].
Enzymatic processing of lignocellulosic biomass: principles, recent advances and perspectives
[J].
DOI:10.1007/s10295-020-02301-8
URL
[本文引用: 1]

Efficient saccharification of lignocellulosic biomass requires concerted development of a pretreatment method, an enzyme cocktail and an enzymatic process, all of which are adapted to the feedstock. Recent years have shown great progress in most aspects of the overall process. In particular, increased insights into the contributions of a wide variety of cellulolytic and hemicellulolytic enzymes have improved the enzymatic processing step and brought down costs. Here, we review major pretreatment technologies and different enzyme process setups and present an in-depth discussion of the various enzyme types that are currently in use. We pay ample attention to the role of the recently discovered lytic polysaccharide monooxygenases (LPMOs), which have led to renewed interest in the role of redox enzyme systems in lignocellulose processing. Better understanding of the interplay between the various enzyme types, as they may occur in a commercial enzyme cocktail, is likely key to further process improvements.
Isolation and characterization of a novel laccase for lignin degradation, LacZ1
[J].
DOI:10.1128/AEM.01355-21
URL
[本文引用: 1]
Lignin is a natural aromatic polymer that is not easily degraded, hindering the efficient use of lignocellulose-rich biomass resources, such as straw. Biodegradation is a method of decomposing lignin that has recently received increasing attention.
Pretreatment of lignocellulosic materials as substrates for fermentation processes
[J].
DOI:10.3390/molecules23112937
URL
[本文引用: 1]
Lignocellulosic biomass is an abundant and renewable resource that potentially contains large amounts of energy. It is an interesting alternative for fossil fuels, allowing the production of biofuels and other organic compounds. In this paper, a review devoted to the processing of lignocellulosic materials as substrates for fermentation processes is presented. The review focuses on physical, chemical, physicochemical, enzymatic, and microbiologic methods of biomass pretreatment. In addition to the evaluation of the mentioned methods, the aim of the paper is to understand the possibilities of the biomass pretreatment and their influence on the efficiency of biofuels and organic compounds production. The effects of different pretreatment methods on the lignocellulosic biomass structure are described along with a discussion of the benefits and drawbacks of each method, including the potential generation of inhibitory compounds for enzymatic hydrolysis, the effect on cellulose digestibility, the generation of compounds that are toxic for the environment, and energy and economic demand. The results of the investigations imply that only the stepwise pretreatment procedure may ensure effective fermentation of the lignocellulosic biomass. Pretreatment step is still a challenge for obtaining cost-effective and competitive technology for large-scale conversion of lignocellulosic biomass into fermentable sugars with low inhibitory concentration.
Synergistic action between extracellular products from white-rot fungus and cellulase significantly improves enzymatic hydrolysis
[J].
DOI:10.1080/21655979.2017.1308991
PMID:28384075
[本文引用: 1]
With a set of perfect extracellular lignocellulolytic enzymes, white-rot fungus has been recognized as playing an important role in the degradation of lignocellulose materials, which leads to the possibility of creating a composite enzymatic system with high hydrolysis efficiency in vitro. Echinodontium taxodii is a promising white-rot fungus for biologic pretreatment. In this study, we extracted the extracellular products of E. taxodii under solid-state fermentation conditions, mixed the extracellular products with cellulase to build a composite enzymatic system, and systematically evaluated the effect of this system on the hydrolysis of acid-pretreated and raw maize stovers. We found that the extracellular products from E. taxodii could significantly improve the hydrolysis efficiency of cellulase, with a synergistic action between the extracellular products and cellulase. Corn stovers treated with extracellular products were suitable for the enzymatic hydrolysis of cellulase. Furthermore, we found that pure proteins from the extracellular products were not sufficient to generate synergistic action. This finding suggests that non-protein substances may also be involved in the synergistic action between the extracellular products and cellulase.
草菇GH61家族基因的生物信息学分析及金属离子对其表达水平的影响
[J].
Bioinformatic and gene expression analyses of the GH61 family genes of Volvariella volvacea
[J].
Oxidized product profiles of AA9 lytic polysaccharide monooxygenases depend on the type of cellulose
[J].
DOI:10.1021/acssuschemeng.1c04100
PMID:34722005
[本文引用: 1]
Lytic polysaccharide monooxygenases (LPMOs) are essential for enzymatic conversion of lignocellulose-rich biomass in the context of biofuels and platform chemicals production. Considerable insight into the mode of action of LPMOs has been obtained, but research on the cellulose specificity of these enzymes is still limited. Hence, we studied the product profiles of four fungal Auxiliary Activity family 9 (AA9) LPMOs during their oxidative cleavage of three types of cellulose: bacterial cellulose (BC), Avicel PH-101 (AVI), and regenerated amorphous cellulose (RAC). We observed that attachment of a carbohydrate-binding module 1 (CBM1) did not change the substrate specificity of LPMO9B from C1 (LPMO9B) but stimulated the degradation of all three types of cellulose. A detailed quantification of oxidized ends in both soluble and insoluble fractions, as well as characterization of oxidized cello-oligosaccharide patterns, suggested that LPMO9B generates mainly oxidized cellobiose from BC, while producing oxidized cello-oligosaccharides from AVI and RAC ranged more randomly from DP2-8. Comparable product profiles, resulting from BC, AVI, and RAC oxidation, were found for three other AA9 LPMOs. These distinct cleavage profiles highlight cellulose specificity rather than an LPMO-dependent mechanism and may further reflect that the product profiles of AA9 LPMOs are modulated by different cellulose types.© 2021 The Authors. Published by American Chemical Society.
Efficient constitutive expression of cellulolytic enzymes in Penicillium oxalicum for improved efficiency of lignocellulose degradation
[J].DOI:10.4014/jmb.2101.01003 URL [本文引用: 1]
An oxidative enzyme boosting the enzymatic conversion of recalcitrant polysaccharides
[J].
DOI:10.1126/science.1192231
PMID:20929773
[本文引用: 1]
Efficient enzymatic conversion of crystalline polysaccharides is crucial for an economically and environmentally sustainable bioeconomy but remains unfavorably inefficient. We describe an enzyme that acts on the surface of crystalline chitin, where it introduces chain breaks and generates oxidized chain ends, thus promoting further degradation by chitinases. This enzymatic activity was discovered and further characterized by using mass spectrometry and chromatographic separation methods to detect oxidized products generated in the absence or presence of H(2)(18)O or (18)O(2). There are strong indications that similar enzymes exist that work on cellulose. Our findings not only demonstrate the existence of a hitherto unknown enzyme activity but also provide new avenues toward more efficient enzymatic conversion of biomass.
Fungal cellulose degradation by oxidative enzymes: from dysfunctional GH61 family to powerful lytic polysaccharide monooxygenase family
[J].
DOI:10.1093/bfgp/elu032
PMID:25217478
[本文引用: 1]
Our understanding of fungal cellulose degradation has shifted dramatically in the past few years with the characterization of a new class of secreted enzymes, the lytic polysaccharide monooxygenases (LPMO). After a period of intense research covering structural, biochemical, theoretical and evolutionary aspects, we have a picture of them as wedge-like copper-dependent metalloenzymes that on reduction generate a radical copper-oxyl species, which cleaves mainly crystalline cellulose. The main biological function lies in the synergism of fungal LPMOs with canonical hydrolytic cellulases in achieving efficient cellulose degradation. Their important role in cellulose degradation is highlighted by the wide distribution and often numerous occurrences in the genomes of almost all plant cell-wall degrading fungi. In this review, we provide an overview of the latest achievements in LPMO research and consider the open questions and challenges that undoubtedly will continue to stimulate interest in this new and exciting group of enzymes. © The Author 2014. Published by Oxford University Press.
A comparative study on the activity of fungal lytic polysaccharide monooxygenases for the depolymerization of cellulose in soybean spent flakes
[J].DOI:10.1016/j.carres.2017.07.004 URL [本文引用: 1]
Functional characterization of cellulose-degrading AA9 lytic polysaccharide monooxygenases and their potential exploitation
[J].
DOI:10.1007/s00253-020-10467-5
PMID:32076777
[本文引用: 1]
Cellulose-degrading auxiliary activity family 9 (AA9) lytic polysaccharide monooxygenases (LPMOs) are known to be widely distributed among filamentous fungi and participate in the degradation of lignocellulose via the oxidative cleavage of celluloses, cello-oligosaccharides, or hemicelluloses. AA9 LPMOs have been reported to have extensive interactions with not only cellulases but also oxidases. The addition of AA9 LPMOs can greatly reduce the amount of cellulase needed for saccharification and increase the yield of glucose. The discovery of AA9 LPMOs has greatly changed our understanding of how fungi degrade cellulose. In this review, apart from summarizing the recent discoveries related to their catalytic reaction, functional diversity, and practical applications, the stability, expression system, and protein engineering of AA9 LPMOs are reviewed for the first time. This review may provide a reference value to further broaden the substrate range of AA9 LPMOs, expand the scope of their practical applications, and realize their customization for industrial utilization.Key Points• The stability and expression system of AA9 LPMOs are reviewed for the first time.• The protein engineering of AA9 LPMOs is systematically summarized for the first time.• The latest research results on the catalytic mechanism of AA9 LPMOs are summarized.• The application of AA9 LPMOs and their relationship with other enzymes are reviewed.
Structure of a C1/C4-oxidizing AA9 lytic polysaccharide monooxygenase from the thermophilic fungus Malbranchea cinnamomea
[J].
DOI:10.1107/S2059798321006628
URL
[本文引用: 1]
The thermophilic fungus Malbranchea cinnamomea contains a host of enzymes that enable its ability as an efficient degrader of plant biomass and that could be mined for industrial applications. This thermophilic fungus has been studied and found to encode eight lytic polysaccharide monooxygenases (LPMOs) from auxiliary activity family 9 (AA9), which collectively possess different substrate specificities for a range of plant cell-wall-related polysaccharides and oligosaccharides. To gain greater insight into the molecular determinants defining the different specificities, structural studies were pursued and the structure of McAA9F was determined. The enzyme contains the immunoglobulin-like fold typical of previously solved AA9 LPMO structures, but contains prominent differences in the loop regions found on the surface of the substrate-binding site. Most significantly, McAA9F has a broad substrate specificity, with activity on both crystalline and soluble polysaccharides. Moreover, it contains a small loop in a region where a large loop has been proposed to govern specificity towards oligosaccharides. The presence of the small loop leads to a considerably flatter and more open surface that is likely to enable the broad specificity of the enzyme. The enzyme contains a succinimide residue substitution, arising from intramolecular cyclization of Asp10, at a position where several homologous members contain an equivalent residue but cyclization has not previously been observed. This first structure of an AA9 LPMO from M. cinnamomea aids both the understanding of this family of enzymes and the exploration of the repertoire of industrially relevant lignocellulolytic enzymes from this fungus.
Novel factors contributing to fungal pathogenicity at early stages of Setosphaeria turcica infection
[J].DOI:10.1111/mpp.13140 URL [本文引用: 1]
来自Yeosuana marina sp. JLT21内切型海藻酸裂解酶的异源表达及酶学表征
[J].
DOI:10.13560/j.cnki.biotech.bull.1985.2021-0539
[本文引用: 1]
褐藻寡糖有着丰富的生物学功能,酶法制备功能性褐藻寡糖具有重要实践应用价值。为发掘高活性及稳定性的褐藻寡糖制备酶,对浅海热液嗜热菌Yeosuana marina sp. JLT21中的海藻酸裂解酶YMA-1的基因在大肠杆菌中进行表达、纯化及酶活鉴定。结果发现YMA-1由306个氨基酸残基构成,是多糖裂解酶家族7(PL7)新成员;重组YMA-1酶的最适催化条件是55℃,pH 9.0,比活力1.3×10<sup>4</sup>U/mg,Cu<sup>2+</sup> 可有效促进酶活;在37℃,pH 9.0 条件下,该酶对海藻酸钠、聚甘露糖醛酸和聚古罗糖醛酸的比活力分别达到(5 201.21±86.46)U/mg、(6 399.73±253.12)U/mg和(3 751.68±116.25)U/mg,酶解海藻酸钠终产物多为不饱和三糖和四糖,表现出内切双功能型海藻酸裂解酶活性。YMA-1酶作为PL7家族中较宽底物谱、高活性及稳定性的内切海藻酸裂解酶,在高效绿色生产功能性褐藻寡糖上有着潜在应用价值。
Heterologous expression and characterization of endo-type alginate lyase from Yeosuana marina sp. JLT21
[J].
过表达Spt7对黑曲霉生长及抗逆性影响
[J].
DOI:10.13560/j.cnki.biotech.bull.1985.2021-0946
[本文引用: 1]
SAGA复合体是一种多功能蛋白复合物,负责细胞内10%以上的基因转录。Spt7作为SAGA复合体的核心蛋白,维持SAGA复合体的稳定。除酵母之外的真菌中并未有Spt7相关的研究报道。本研究对不同黑曲霉、丝衣霉等丝状真菌来源的Spt7的氨基酸序列进行多序列比对及蛋白结构的分析,发现Spt7在不同物种一致性较低,但均存在保守型的Bromo结构域。以黑曲霉1062为出发菌株,通过农杆菌转化法将过表达spt7的质粒转入1062菌株中,获得黑曲霉OE spt7转化子。通过对OE spt7转化子及对照组在CM培养基生长形态进行观察分析发现,过表达Spt7有益于菌体的生长及分生孢子的生成,在72 h时OE spt7转化子和对照组的菌落直径分别达到2.9 cm和2.8 cm,且转化子的孢子数量达到(5.8-6.3)×10<sup>7</sup>/cm<sup>2</sup>,较对照组(2.3×10<sup>7</sup>/cm<sup>2</sup>)提高1.5-1.7倍,且转化子菌丝分支较对照组多。另外,与对照组相比,OE spt7转化子在15 mmol/L H<sub>2</sub>O<sub>2</sub>条件下孢子萌发更快,菌落直径是对照组4倍,且在15% NaCl高渗以及39℃高温的条件下较对照组生长更加鲜活。通过qRT-PCR分析参与菌体抗氧化胁迫的过氧化物酶及耐高温的热休克蛋白基因的转录水平,发现除了CatR转录水平下调3.56倍,其他的过氧化物酶SOD、CpeB、GPX分别上调了3.8、1.89、3.56倍,除Hsp40及Hsp70较对照组无显著差异,Hsp90的转录水平上调19倍。
Effect of Spt7 overexpression of on the growth and stress resistance of Aspergillus niger
[J].
A fast and sensitive activity assay for lytic polysaccharide monooxygenase
[J].
DOI:10.1186/s13068-018-1063-6
PMID:29588664
[本文引用: 1]
Background: Lytic polysaccharide monooxygenases (LPMO) release a spectrum of cleavage products from their polymeric substrates cellulose, hemicellulose, or chitin. The correct identification and quantitation of these released products is the basis of MS/HPLC-based detection methods for LPMO activity. The duration, effort, and intricate analysis allow only specialized laboratories to measure LPMO activity in day-to-day work. A spectrophotometric assay will simplify the screening for LPMO in culture supernatants, help monitor recombinant LPMO expression and purification, and support enzyme characterization.Results: Based on a newly discovered peroxidase activity of LPMO, we propose a fast, robust, and sensitive spectrophotometric activity assay using 2,6-dimethoxyphenol (2,6-DMP) and H2O2. The fast enzymatic assay (300 s) consists of 1 mM 2,6-DMP as chromogenic substrate, 100 mu M H2O2 as cosubstrate, and an adequate activity of LPMO in a suitable buffer. The high molar absorption coefficient of the formed product coerulignone (epsilon(469) = 53,200 M-1 cm(-1)) makes the assay sensitive and allows reliable activity measurements of LPMO in concentrations of approx. 0.5-50 mg L-1.Conclusions: The activity assay based on 2,6-DMP detects a novel peroxidase activity of LPMO. This activity can be accurately measured and used for enzyme screening, production, and purification, and can also be applied to study binding constants or thermal stability. However, the assay has to be used with care in crude extracts, because other enzymes such as laccase or peroxidase will interfere with the assay. We also want to stress that the peroxidase activity is a homogeneous reaction with soluble substrates and should not be correlated to heterogeneous LPMO activity on polymeric substrates.
Transcriptome and secretome analysis of Aspergillus fumigatus in the presence of sugarcane bagasse
[J].DOI:10.1186/s12864-018-4627-8 [本文引用: 1]
丝状真菌溶解性多糖单加氧酶的研究进展
[J].
Research progress on lytic polysaccharide monooxygenases(LPMOs)in filamentous fungi
[J].
Analysis of the transcriptome in Aspergillus tamarii during enzymatic degradation of sugarcane bagasse
[J].DOI:10.3389/fbioe.2018.00123 URL [本文引用: 1]
Lytic polysaccharide monooxygenase from Aspergillus fumigatus can improve enzymatic cocktail activity during sugarcane bagasse hydrolysis
[J].
DOI:10.2174/0929866526666190228163629
URL
[本文引用: 1]
Lytic Polysaccharide Monooxygenases (LPMOs) are auxiliary accessory\nenzymes that act synergistically with cellulases and which are increasingly being used in secondgeneration\nbioethanol production from biomasses. Several LPMOs have been identified in various\nfilamentous fungi, including Aspergillus fumigatus. However, many LPMOs have not been characterized\nyet.
大肠杆菌分泌表达裂解性多糖单加氧酶发酵条件的优化
[J].
Fermentation condition optimization of recombinant lytic polysaccharide monooxygenase extracellularly expressed in Escherichia coli
[J].
LPMO af AA9_B and cellobiohydrolase af Cel6A from A. fumigatus boost enzymatic saccharification activity of cellulase cocktail
[J].
DOI:10.3390/ijms22010276
URL
[本文引用: 1]
Cellulose is the most abundant polysaccharide in lignocellulosic biomass, where it is interlinked with lignin and hemicellulose. Bioethanol can be produced from biomass. Since breaking down biomass is difficult, cellulose-active enzymes secreted by filamentous fungi play an important role in degrading recalcitrant lignocellulosic biomass. We characterized a cellobiohydrolase (AfCel6A) and lytic polysaccharide monooxygenase LPMO (AfAA9_B) from Aspergillus fumigatus after they were expressed in Pichia pastoris and purified. The biochemical parameters suggested that the enzymes were stable; the optimal temperature was ~60 °C. Further characterization revealed high turnover numbers (kcat of 147.9 s−1 and 0.64 s−1, respectively). Surprisingly, when combined, AfCel6A and AfAA9_B did not act synergistically. AfCel6A and AfAA9_B association inhibited AfCel6A activity, an outcome that needs to be further investigated. However, AfCel6A or AfAA9_B addition boosted the enzymatic saccharification activity of a cellulase cocktail and the activity of cellulase Af-EGL7. Enzymatic cocktail supplementation with AfCel6A or AfAA9_B boosted the yield of fermentable sugars from complex substrates, especially sugarcane exploded bagasse, by up to 95%. The synergism between the cellulase cocktail and AfAA9_B was enzyme- and substrate-specific, which suggests a specific enzymatic cocktail for each biomass by up to 95%. The synergism between the cellulase cocktail and AfAA9_B was enzyme- and substrate-specific, which suggests a specific enzymatic cocktail for each biomass.
米曲霉裂解性多糖单加氧酶的异源表达与性质分析
[J].
Heterologous expression and characterization of Aspergillus oryzae lytic polysaccharide monooxygenases
[J].
嗜热毛壳菌多糖单加氧酶的氧化特性及协同作用
[J].
The oxidation properties and synergism of polysaccharide monooxygenase from Chaetomium thermophilum
[J].
Thermal unfolding and refolding of a lytic polysaccharide monooxygenase from Thermoascus aurantiacus
[J].DOI:10.1039/C9RA05920B URL [本文引用: 1]
A thermostable bacterial lytic polysaccharide monooxygenase with high operational stability in a wide temperature range
[J].
DOI:10.1186/s13068-020-01834-5
PMID:33292445
[本文引用: 1]
Lytic polysaccharide monooxygenases (LPMOs) are oxidative, copper-dependent enzymes that function as powerful tools in the turnover of various biomasses, including lignocellulosic plant biomass. While LPMOs are considered to be of great importance for biorefineries, little is known about industrial relevant properties such as the ability to operate at high temperatures. Here, we describe a thermostable, cellulose-active LPMO from a high-temperature compost metagenome (called mgLPMO10).MgLPMO10 was found to have the highest apparent melting temperature (83 °C) reported for an LPMO to date, and is catalytically active up to temperatures of at least 80 °C. Generally, mgLPMO10 showed good activity and operational stability over a wide temperature range. The LPMO boosted cellulose saccharification by recombinantly produced GH48 and GH6 cellobiohydrolases derived from the same metagenome, albeit to a minor extent. Cellulose saccharification studies with a commercial cellulase cocktail (Celluclast®) showed that the performance of this thermostable bacterial LPMO is comparable with that of a frequently utilized fungal LPMO from Thermoascus aurantiacus (TaLPMO9A).The high activity and operational stability of mgLPMO10 are of both fundamental and applied interest. The ability of mgLPMO10 to perform oxidative cleavage of cellulose at 80 °C and the clear synergy with Celluclast® make this enzyme an interesting candidate in the development of thermostable enzyme cocktails for use in lignocellulosic biorefineries.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}