







生物技术通报 ›› 2025, Vol. 41 ›› Issue (10): 72-86.doi: 10.13560/j.cnki.biotech.bull.1985.2025-0620
范艳飞1,2,3( ), 叶露幻1,2, 李雨桐1,2,3, 王钏跞1,2,3, 张瑞1,2, 罗建华1,2,3, 王鹏1,2()
), 叶露幻1,2, 李雨桐1,2,3, 王钏跞1,2,3, 张瑞1,2, 罗建华1,2,3, 王鹏1,2()
收稿日期:2025-06-13
出版日期:2025-10-26
发布日期:2025-07-30
通讯作者:
王鹏,研究员,研究方向 :植物光合机构塑造及运转调控;E-mail: wangpeng@cemps.ac.cn作者简介:范艳飞,博士研究生,研究方向 :环式光合电子传递的遗传调控机制;E-mail: fanyanfei@cemps.ac.cn
基金资助:
FAN Yan-fei1,2,3(), YE Lu-huan1,2, LI Yu-tong1,2,3, WANG Chuan-luo1,2,3, ZHANG Rui1,2, LUO Jian-hua1,2,3, WANG Peng1,2()
Received:2025-06-13
Published:2025-10-26
Online:2025-07-30
摘要:
目的 在全球气候变暖以及粮食需求不断增长的背景下,粮食安全问题愈发严峻,要求作物在增产的同时提高环境胁迫适应能力。光合作用是作物产量形成的基础,其光反应阶段的环式光合电子传递途径偶联ATP合成并调节还原力积累,对植物的高温响应和高光效具有重要作用,因此本研究致力于相关遗传调控因子的发掘和应用。 方法 利用具有环式光合电子传递活性差异的代表性小麦品系为参照,结合光合参数测量、蛋白含量测定和转录组测序技术,从小麦小偃54和京411杂交培育的重组自交系后代中筛选获得了具有环式电子传递活性两极分离的株系及其差异表达基因。 结果 其中环式电子传递活性较高的株系同时表现出较高的线性电子传递活性和光合CO2同化速率,并在光强升高条件下保持优势,可作为高光效育种材料加以应用。通过差异表达基因分析挖掘出一系列具有促进环式光合电子传递或整体光合活性潜力的功能基因和转录因子。将部分基因(包括TaPnsL2和TaNAC等)构建过量表达载体转化水稻栽培品种秀水134,T1代和T2代转基因材料在海南省和上海大田试验表现出光合速率升高的优势。 结论 针对环式电子传递活性的遗传筛选或改造,具有提高作物光合效率或强光适应性的潜力。
范艳飞, 叶露幻, 李雨桐, 王钏跞, 张瑞, 罗建华, 王鹏. 运用小麦杂交株系挖掘环式光合电子传递调控基因并应用于作物高光效改造[J]. 生物技术通报, 2025, 41(10): 72-86.
FAN Yan-fei, YE Lu-huan, LI Yu-tong, WANG Chuan-luo, ZHANG Rui, LUO Jian-hua, WANG Peng. Utilizing Wheat Hybrid Lines to Mine Genes Regulating Cyclic Electron Flow and Applying Them in Improving Photosynthetic Efficiency in Crops[J]. Biotechnology Bulletin, 2025, 41(10): 72-86.

图1 小麦重组自交系群体种植及代表性品种的P700+暗还原初始速率和ETR(Ⅰ)-ETR(Ⅱ)测量A: 上方为进行第一轮P700+还原速率测定时其中1批的小麦材料实拍图;左下为5个小麦代表性品种;右下为在大田生长的小麦群体。B: 左侧为5个小麦代表性品种P700+的还原曲线实测图,右侧为对应的曲线下降初始斜率柱状图。C: 5个小麦代表性品种在光强梯度升高情况下的ETR(Ⅰ)-ETR(Ⅱ)折线图
Fig. 1 Planting of wheat recombinant inbred population, and measurements of initial P700+ dark-reduction rates and ETR(Ⅰ)- ETR(Ⅱ) in representative varietiesA: The above is a photo of one batch of wheat materials during the first round of P700+ reduction rate measurement; the bottom left shows five representative wheat varieties; the bottom right displays the wheat population growing in the field. B: Left, P700+reduction curves of five representative wheat varieties; right, initial slope of curve decline (bar chart). C: Line chart of ETR(Ⅰ)-ETR(Ⅱ) for five representative wheat varieties under increasing light intensity gradients
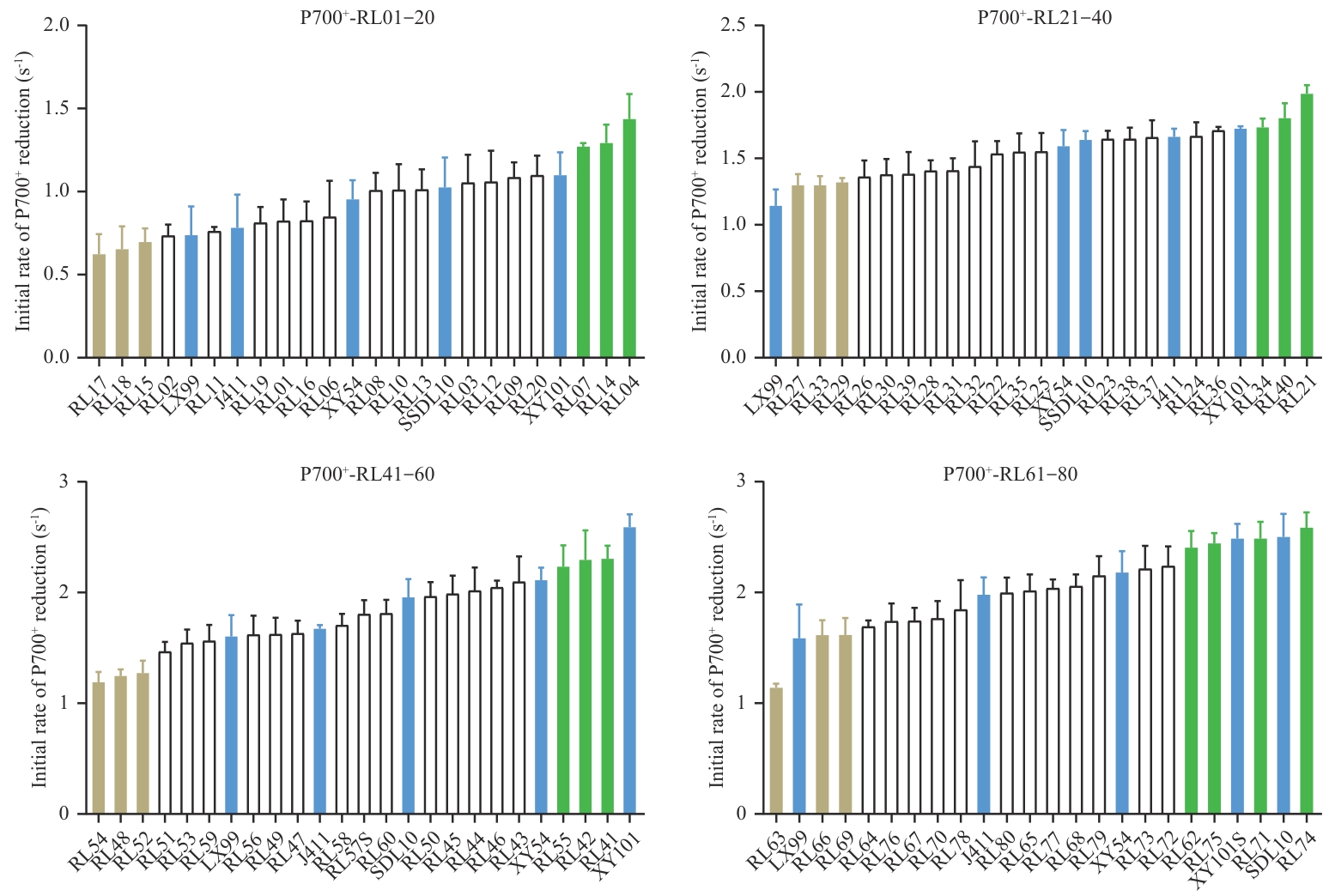
图2 第一轮测定小麦群体P700+暗还原初始速率分布示例第一轮P700+暗还原初始速率筛选覆盖了整个小麦重组自交系群体,此处显示前80个株系(RL01-RL80,“RL”代表重组自交系)的测量结果,其中每连续的20个株系加上5个代表性品种作为一组进行测量和作图。代表性品种以蓝色显示,在数据分布的两端选出环式电子传递活性高和低的株系,分别以绿色和浅棕色显示
Fig. 2 Distribution examples of initial P700+ dark-reduction rates of wheat population in the first-round measurementThe first-round screening of initial P700+ dark-reduction rates covered the entire wheat recombinant inbred line population. Here, the measurements of the first 80 lines (RL01-RL80, “RL” is short for “recombinant inbred line”) are displayed, with every consecutive 20 lines plus 5 representative cultivars measured and plotted as a group. The representative varieties are displayed in blue, while the lines with high and low cyclic electron transport activity selected from both ends of the data distribution are shown in green and light brown, respectively

图3 第二轮测定小麦群体P700+暗还原初始速率分布图第二轮P700+暗还原初始速率筛选面向第一轮筛选出的环式电子传递活性高和低的株系,将其随机编入4个包含代表性品种的批次采用与第一轮相同的方式进行测量和作图。代表性品种仍以蓝色显示,第一轮测定筛选出的环式电子传递活性高和低的株系,仍以绿色和浅棕色显示。“RL”代表重组自交系
Fig. 3 Distribution of initial P700+ dark reduction rates of wheat population in the second-round measurementThe second-round initial P700+ dark-reduction rate screening focused on lines with either high or low cyclic electron transport activity from the. They were randomly grouped into 4 batches alongside representative cultivars and measured and graphed usingthe same protocol as the first-round screening. Representative varieties are still shown in blue, while the lines with high and low cyclic electron transport activity selected from the first-round screening remain displayed in green and light brown, respectively. “RL” is short for “recombinant inbred line”
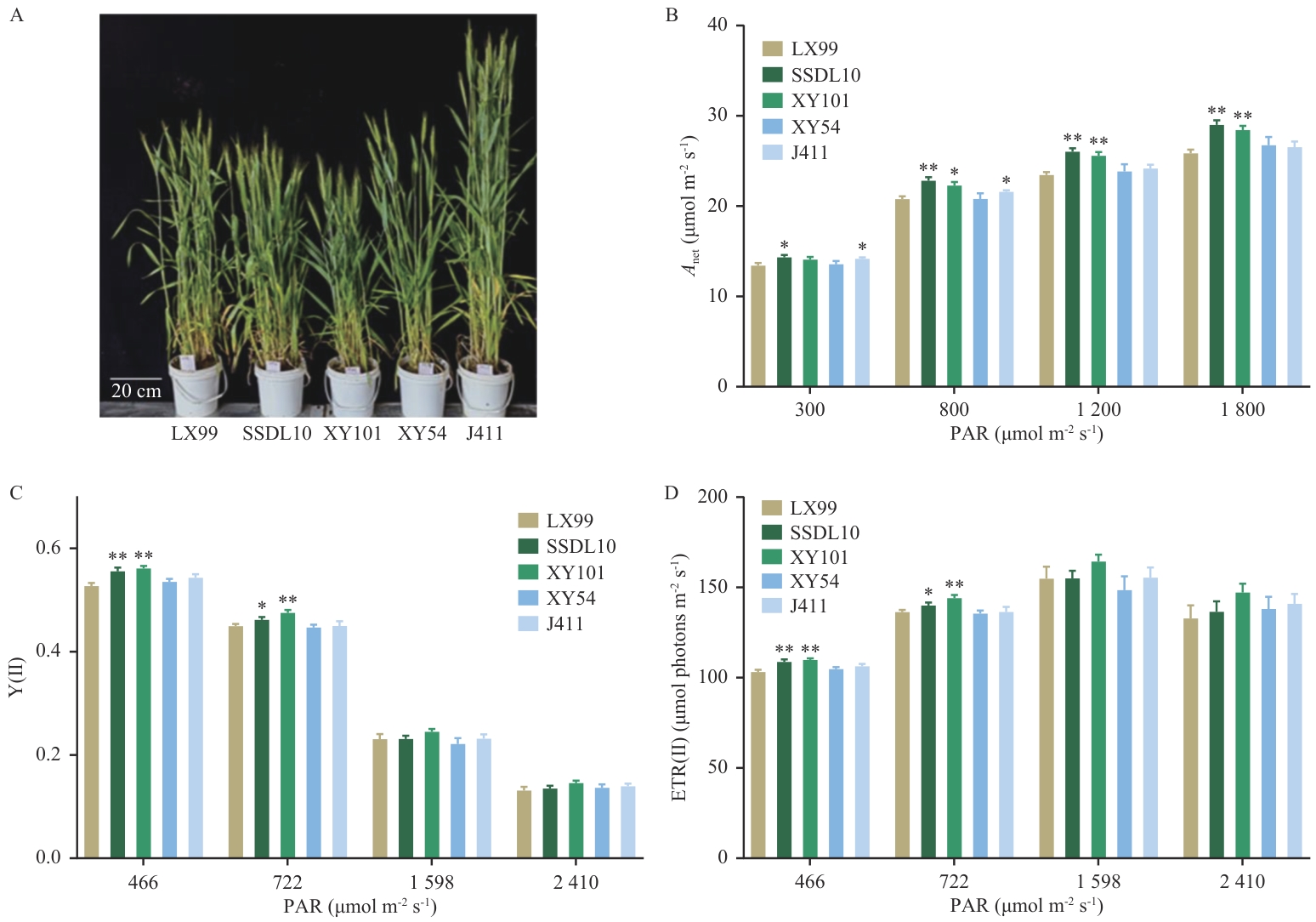
图4 代表性小麦品种的生长表型及在梯度光强下的光合参数A: 抽穗后的5个代表性小麦品种;B-D: 代表性品种在光强梯度下的光合速率Anet、Y(Ⅱ)和ETR(Ⅱ)。大田小麦抽穗后,在晴朗的上午使用Li6400光合仪和PAM2000便携式叶绿素荧光仪测定了小麦代表性品种LX99、SSDL10、XY101、XY54和J411的旗叶在不同光强下的光合速率及电子传递活性,生物学重复为4-6个
Fig. 4 Growth phenotypes of representative wheat cultivars and light-intensity-dependent photosynthetic parametersA: Five representative wheat varieties after heading. B-D: Net photosynthetic rate (Anet), Y(Ⅱ) and ETR(Ⅱ) of representative varieties under light-intensity gradients. Field measurements of photosynthetic rates and electron transport activities under varying light intensities were conducted on flag leaves of wheat representative cultivars (LX99,SSDL10,XY101, XY54, and J411) using Li-6400 photosynthesis system and PAM-2000 portable chlorophyll fluorometer. The measurements were performed on clear mornings during the post-heading stage, with 4-6 biological replicates per cultivar
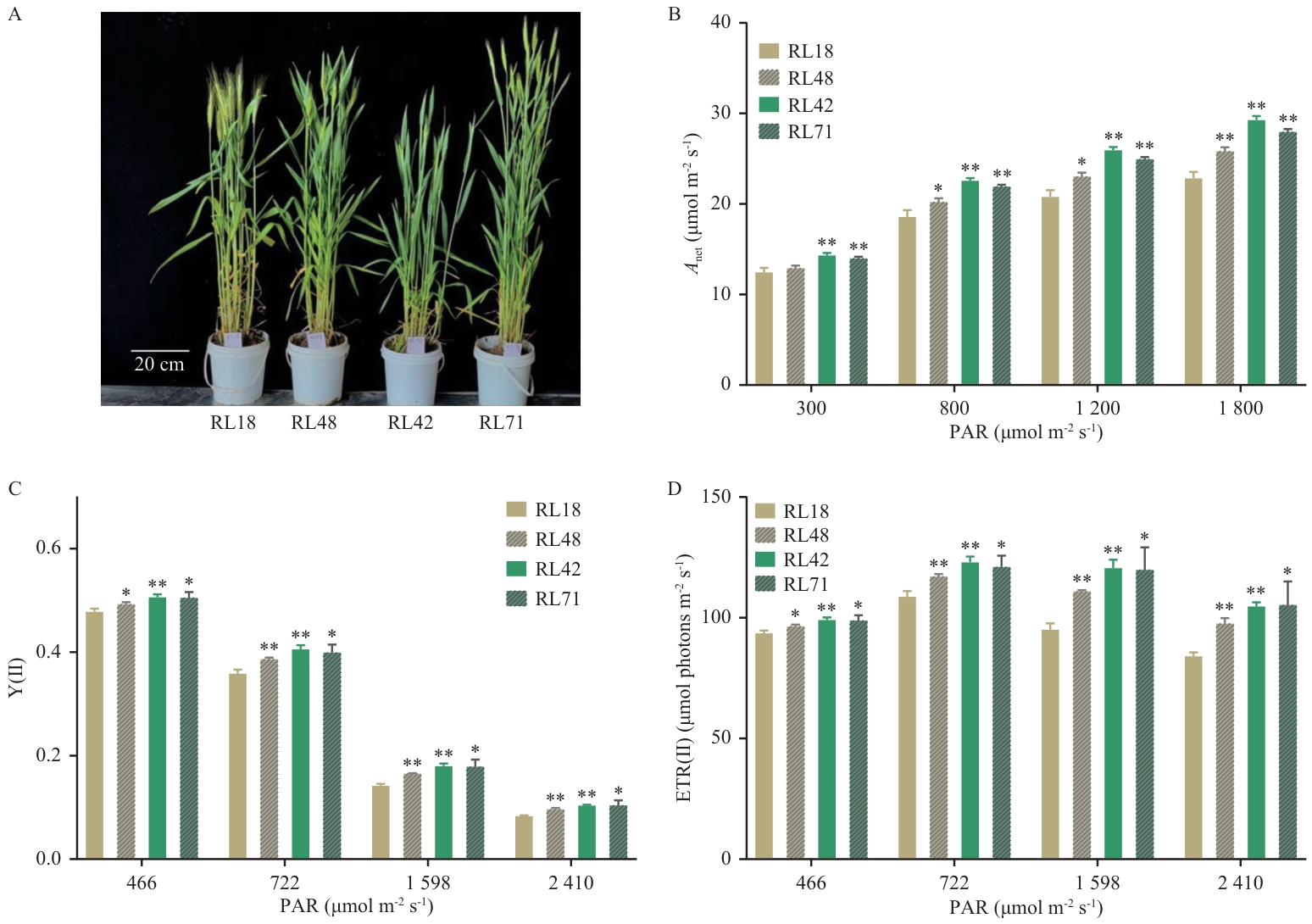
图5 小麦重组自交株系的生长表型及在梯度光强下的光合参数A: 抽穗后的4个小麦重组自交株系;B-D:分别为4个小麦重组自交株系在光强梯度下的光合速率Anet、Y(Ⅱ)和ETR(Ⅱ)。大田小麦抽穗后,在晴朗的上午使用Li6400光合仪和PAM2000便携式叶绿素荧光仪测定了小麦重组自交系群体株系RL18、RL48、RL42和RL71的旗叶在不同光强下的光合速率及电子传递活性,生物学重复为5-6个
Fig. 5 Growth phenotypes of wheat recombinant inbred lines and light-intensity-dependent photosynthetic parametersA: Four wheat recombinant inbred lines after heading. B-D: Net photosynthetic rate (Anet), Y(Ⅱ) and ETR(Ⅱ) of wheat recombinant inbred lines under light-intensity gradients, respectively. Field measurements of photosynthetic rates and electron transport activities under varying light intensities were conducted on flag leaves of wheat recombinant inbred lines (RL18, RL48, RL42 and RL71) using Li-6400 photosynthesis system and PAM-2000 portable chlorophyll fluorometer. The measurements were performed on clear mornings during the post-heading stage, with 5-6 biological replicates per cultivar

图6 重组自交系群体NDH亚基含量测定A: 重组自交系群体NDH-H亚基含量测定(第一轮);B: 重组自交系群体NDH含量第二轮测定。每张免疫杂交(Western blot)图内部具有可比性,杂交图之间显色强度可能不同。每张杂交图均含有XY101(NDH-H含量高)和LX99(NDH-H含量低)作为参考
Fig. 6 Determination of NDH subunit content in recombinant inbred line populationA: Determination of NDH-H subunit content in recombinant inbred line population (first round). B: Second-round quantification of NDH complex content in the recombinant inbred line population. The Western blot results are comparable within each blot, but the signal intensities may vary between different blots. Each immunoblotting image contains XY101 (high NDH-H content) and LX99 (low NDH-H content) as references

图7 转录组测序结果分析示例A-D: 分别显示光系统间电子递体、环式光合电子传递、光系统I和转录因子相关基因差异表达。热图显示各品种FPKM数值相对于“LX99+J411”平均值的倍数的对数值|Log2(FC)|
Fig. 7 Analysis example of transcriptome sequencing resultsA-D: Differential expressions of genes related to intersystem electron carriers, cyclic photosynthetic electron transport, PSI and transcription factors respectively. The heatmap demonstrates the logarithmic values |Log2(FC)| of the fold change of FPKM values for each variety relative to the average of “LX99+J411”

图8 小麦PnsL2和NAC基因过量表达的T1和T2代水稻株系光合速率分析A-C: 小麦PnsL2基因过量表达水稻于海南和上海、不同测量光强下的净光合速率(Anet);D-F:小麦转录因子基因NAC过量表达水稻于海南和上海、不同测量光强下的净光合速率(Anet)
Fig. 8 Photosynthetic rate analysis of T1 and T2 generation rice lines overexpressing wheat PnsL2 and NAC genesA-C: Net photosynthetic rate (Anet) of wheat PnsL2 gene-overexpressing rice lines under different measuring light intensities in Hainan and Shanghai. D-F: Net photosynthetic rate (Anet) of wheat transcription factor gene NAC-overexpressing rice lines under different measuring light intensities in Hainan and Shanghai
| [1] | McGuire S. FAO, IFAD, and WFP. the state of food insecurity in the world 2015: meeting the 2015 international hunger targets: taking stock of uneven progress. Rome: FAO, 2015 [J]. Adv Nutr, 2015, 6(5): 623-624. |
| [2] | Mazza JJ. Climate change and agriculture: future implications [J]. WMJ, 2017, 116(4): 191. |
| [3] | Nakamura N, Iwano M, Havaux M, et al. Promotion of cyclic electron transport around photosystem I during the evolution of NADP-malic enzyme-type C4 photosynthesis in the genus Flaveria [J]. New Phytol, 2013, 199(3): 832-842. |
| [4] | Zhu X-G, Long SP, Ort DR. Improving photosynthetic efficiency for greater yield [J]. Annu Rev Plant Biol, 2010, 61: 235-261. |
| [5] | Bailey-Serres J, Parker JE, Ainsworth EA, et al. Genetic strategies for improving crop yields [J]. Nature, 2019, 575(7781): 109-118. |
| [6] | Ma MZ, Liu YF, Bai CM, et al. The significance of chloroplast NAD(P)H dehydrogenase complex and its dependent cyclic electron transport in photosynthesis [J]. Front Plant Sci, 2021, 12: 661863. |
| [7] | Yamori W, Makino A, Shikanai T. A physiological role of cyclic electron transport around photosystem Ⅰ in sustaining photosynthesis under fluctuating light in rice [J]. Sci Rep, 2016, 6: 20147. |
| [8] | Shen LL, Tang KL, Wang WD, et al. Architecture of the chloroplast PSI-NDH super complex in Hordeum vulgare . [J]. Nature, 2022, 601(7894): 649-654. |
| [9] | Lu JZ, Yin ZP, Lu T, et al. Cyclic electron flow modulate the linear electron flow and reactive oxygen species in tomato leaves under high temperature [J]. Plant Sci, 2020, 292: 110387. |
| [10] | Sun YJ, Geng QW, Du YP, et al. Induction of cyclic electron flow around photosystem I during heat stress in grape leaves [J]. Plant Sci, 2017, 256: 65-71. |
| [11] | Yamori W, Sakata N, Suzuki Y, et al. Cyclic electron flow around photosystem Ⅰ via chloroplast NAD(P)H dehydrogenase (NDH) complex performs a significant physiological role during photosynthesis and plant growth at low temperature in rice [J]. Plant J, 2011, 68(6): 966-976. |
| [12] | Yamori W, Shikanai T, Makino A. Photosystem I cyclic electron flow via chloroplast NADH dehydrogenase-like complex performs a physiological role for photosynthesis at low light [J]. Sci Rep, 2015, 5: 13908. |
| [13] | Yamori W, Shikanai T. Physiological functions of cyclic electron transport around photosystem I in sustaining photosynthesis and plant growth [J]. Annu Rev Plant Biol, 2016, 67: 81-106. |
| [14] | Jia HS, Oguchi R, Hope AB, et al. Differential effects of severe water stress on linear and cyclic electron fluxes through Photosystem Ⅰ in spinach leaf discs in CO2-enriched air [J]. Planta, 2008, 228(5): 803-812. |
| [15] | Kohzuma K, Cruz JA, Akashi K, et al. The long-term responses of the photosynthetic proton circuit to drought [J]. Plant Cell Environ, 2009, 32(3): 209-219. |
| [16] | Strand DD, Livingston AK, Satoh-Cruz M, et al. Activation of cyclic electron flow by hydrogen peroxide in vivo [J]. Proc Natl Acad Sci U S A, 2015, 112(17): 5539-5544. |
| [17] | Joliot P, Joliot A. Cyclic electron transfer in plant leaf [J]. Proc Natl Acad Sci U S A, 2002, 99(15): 10209-10214. |
| [18] | Breyton C, Nandha B, Johnson GN, et al. Redox modulation of cyclic electron flow around photosystem Ⅰ in C3 plants [J]. Biochemistry, 2006, 45(45): 13465-13475. |
| [19] | Livingston AK, Kanazawa A, Cruz JA, et al. Regulation of cyclic electron flow in C3 plants: differential effects of limiting photosynthesis at ribulose-1, 5-bisphosphate carboxylase/oxygenase and glyceraldehyde-3-phosphate dehydrogenase [J]. Plant Cell Environ, 2010, 33(11): 1779-1788. |
| [20] | Wang HW, Su JH, Shen YG. Difference in response of photosynthesis to bisulfite between two wheat genotypes [J]. J Plant Physiol Mol Biol, 2003, 29(1): 27-32. |
| [21] | 程建峰, 马为民, 陈根云, 等. 小偃54和京411及其杂交后代稳定优选株系光合特性的动态变化 [J]. 作物学报, 2009, 35(6): 1051-1058. |
| Cheng JF, Ma WM, Chen GY, et al. Dynamic changes of photosynthetic characteristics in Xiaoyan 54, Jing 411, and the stable selected superior strains of their hybrid progenies [J]. Acta Agron Sin, 2009, 35(6): 1051-1058. | |
| [22] | He ZH, Li HW, Shen YK, et al. Comparative analysis of the chloroplast proteomes of a wheat (Triticum aestivum L.) single seed descent line and its parents [J]. J Plant Physiol, 2013, 170(13): 1139-1147. |
| [23] | 魏家绵, 沈允钢, 李德耀, 等. 亚硫酸氢钠在低光强下对叶绿体循环光合磷酸化的促进作用 [J]. 植物生理学报, 1989, 15(1): 101-104. |
| Wei JM, Shen YG, Li DY, et al. Stimulatory effect of sodium bisulfite on cyclic-photophosphorylation of chloroplasts under low light intensity [J]. Plant Physiol J, 1989, 15(1): 101-104. | |
| [24] | 王宏炜, 魏家绵, 沈允钢, 等. 低浓度NaHSO3促进田间水稻的光合磷酸化和光合作用(英文) [J]. 植物学报, 2000, 12: 1295-1299. |
| Wang HW, Wei JM, Shen RG, et al. Enhancement of photophosphorylation and photosynthesis in rice by low concentrations of NaHSO3: Under field conditions [J]. J Integr Plant Biol, 2000, (12): 1295-1299. | |
| [25] | Wu YX, Zheng FF, Ma WM, et al. Regulation of NAD(P)H dehydrogenase-dependent cyclic electron transport around PSI by NaHSO3 at low concentrations in tobacco chloroplasts [J]. Plant Cell Physiol, 2011, 52(10): 1734-1743. |
| [26] | 李娜, 韦嘉励, 李庆华, 等. 低浓度NaHSO3促进拟南芥PGR5/PGRL1介导的循环电子传递途径的运转 [J]. 植物生理学报, 2016, 52(11): 1745-1751. |
| Li N, Wei JL, Li QH, et al. Low concentration of NaHSO3 enhances the cyclic electron transport pathway mediated by PGR5/PGRL1 in Arabidopsis thaliana [J]. Plant Physiol J, 2016, 52(11): 1745-1751. | |
| [27] | Hashimoto M, Endo T, Peltier G, et al. A nucleus-encoded factor, CRR2, is essential for the expression of chloroplast ndhB in Arabidopsis [J]. Plant J, 2003, 36(4): 541-549. |
| [28] | Kotera E, Tasaka M, Shikanai T. A pentatricopeptide repeat protein is essential for RNA editing in chloroplasts [J]. Nature, 2005, 433(7023): 326-330. |
| [29] | Peng LW, Cai WH, Shikanai T. Retracted: Chloroplast stromal proteins, CRR6 and CRR7, are required for assembly of the NAD(P)H dehydrogenase sub complex A in Arabidopsis [J]. Plant J, 2010, 63(2): 203-211. |
| [30] | Long TA, Okegawa Y, Shikanai T, et al. Conserved role of proton gradient regulation 5 in the regulation of psi cyclic electron transport [J]. Planta, 2008, 228(6): 907-918. |
| [31] | Okegawa Y, Long TA, Iwano M, et al. A balanced PGR5 level is required for chloroplast development and optimum operation of cyclic electron transport around photosystem I [J]. Plant Cell Physiol, 2007, 48(10): 1462-1471. |
| [32] | Tazoe Y, Ishikawa N, Shikanai T, et al. Overproduction of PGR5 enhances the electron sink downstream of photosystem I in a C4 plant, Flaveria bidentis . [J]. Plant J, 2020, 103(2): 814-823. |
| [33] | Ishikawa N, Yokoe Y, Nishimura T, et al. PsbQ-like protein 3 functions as an assembly factor for the chloroplast NADH dehydrogenase-like complex in Arabidopsis [J]. Plant Cell Physiol, 2020, 61(7): 1252-1261. |
| [34] | Xiong HY, He HD, Chang Y, et al. Multiple roles of NAC transcription factors in plant development and stress responses [J]. J Integr Plant Biol, 2025, 67(3): 510-538. |
| [35] | Ma B, Zhang Y, Fan YF, et al. Genetic improvement of phosphate-limited photosynthesis for high yield in rice [J]. Proc Natl Acad Sci U S A, 2024, 121(34): e2404199121. |
| [1] | 王芳, 邵会茹, 吕林龙, 赵点, 胡振, 吕建珍, 姜亮. 植物和细菌TurboID邻近蛋白标记方法的建立[J]. 生物技术通报, 2025, 41(9): 44-53. |
| [2] | 邓美壁, 严浪, 詹志田, 朱敏, 和玉兵. RUBY辅助的水稻高效CRISPR基因编辑[J]. 生物技术通报, 2025, 41(8): 65-73. |
| [3] | 程雪, 付颖, 柴晓娇, 王红艳, 邓欣. 谷子LHC基因家族鉴定及非生物胁迫表达分析[J]. 生物技术通报, 2025, 41(8): 102-114. |
| [4] | 豆飞飞, 任毓昭, 王石磊, 刘春颖, 王晓东, 王昭懿, 刘彩霞, 刘凤楼, 王掌军, 李清峰. 宁春4号小麦EMS突变体库的构建及表型变异分析[J]. 生物技术通报, 2025, 41(8): 92-101. |
| [5] | 李成花, 豆飞飞, 任毓昭, 刘彩霞, 刘凤楼, 王掌军, 李清峰. 外施水杨酸对白粉菌侵染小麦的影响及白粉菌转录组分析[J]. 生物技术通报, 2025, 41(7): 272-280. |
| [6] | 侯鹰翔, 费思恬, 黎妮, 李兰, 宋松泉, 王伟平, 张超. 水稻miRNAs响应生物胁迫研究进展[J]. 生物技术通报, 2025, 41(7): 69-80. |
| [7] | 张越, 毕钰, 慕雪男, 郑子薇, 王志刚, 徐伟慧. 小麦赤霉病拮抗菌JB7的生防特性[J]. 生物技术通报, 2025, 41(7): 261-271. |
| [8] | 吴浩, 董伟峰, 贺子天, 李艳肖, 谢辉, 孙明哲, 沈阳, 孙晓丽. 水稻BXL基因家族的全基因组鉴定及表达分析[J]. 生物技术通报, 2025, 41(6): 87-98. |
| [9] | 刘园园, 陈析丰, 钱前, 高振宇. 水稻穗发育调控的分子机制研究进展[J]. 生物技术通报, 2025, 41(5): 1-13. |
| [10] | 王轶民, 李莹, 董海涛, 张恒瑞, 常璐, 高田甜, 韩德俊, 吴建辉. SRO家族蛋白在小麦多倍化进程中的演化规律[J]. 生物技术通报, 2025, 41(5): 70-81. |
| [11] | 杜量衡, 唐黄磊, 张治国. 控制水稻光响应基因ELM1的图位克隆[J]. 生物技术通报, 2025, 41(5): 82-89. |
| [12] | 王伟伟, 赵振杰, 王志, 邹景伟, 罗政辉, 张玉杰, 钮力亚, 于亮, 杨学举. 盐胁迫下与小麦生理响应相关的耐盐基因研究进展[J]. 生物技术通报, 2025, 41(5): 14-22. |
| [13] | 陈晓军, 惠建, 马洪文, 白海波, 钟楠, 李稼润, 樊云芳. 利用单碱基基因编辑技术创制OsALS抗除草剂水稻种质资源[J]. 生物技术通报, 2025, 41(4): 106-114. |
| [14] | 王斌, 王玉昆, 肖艳辉. 丁香罗勒(Ocimum gratissimum)叶片响应镉胁迫的比较转录组学分析[J]. 生物技术通报, 2025, 41(3): 255-270. |
| [15] | 马耀武, 张麒宇, 杨淼, 蒋诚, 张振宇, 张伊琳, 李梦莎, 许嘉阳, 张斌, 崔光周, 姜瑛. 烟草根际促生菌的筛选鉴定及促生性能研究[J]. 生物技术通报, 2025, 41(3): 271-281. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||